Số điện thoại: 024 6683 9670
Đăng ký
Đăng nhập
Vi khuẩn trong tiếng Anh là “bacterium”, viết số nhiều là “bacteria”, là cách viết Latinh của tiếng Hy Lạp Cổ đại “βακτήριον”, có nghĩa là "cây gậy, cây gậy", vì những vi khuẩn đầu tiên được phát hiện có hình que.
Vi khuẩn là một nhóm vi sinh vật nhân sơ đơn bào có kích thước rất nhỏ, đôi khi còn được gọi là vi trùng, một số thuộc loại ký sinh trùng. Chúng tạo thành một vùng lớn các vi sinh vật nhân sơ (prokaryotic). Có chiều dài điển hình là vài micromet, vi khuẩn là một trong những dạng sống đầu tiên xuất hiện trên Trái đất và có mặt trong hầu hết các môi trường sống của nó.
Vi khuẩn sống trong đất, nước, suối nước nóng có tính axit, chất thải phóng xạ, và sinh quyển sâu của vỏ Trái đất. Vi khuẩn rất quan trọng trong nhiều giai đoạn của chu trình dinh dưỡng bằng cách tái chế các chất dinh dưỡng như cố định nitơ từ khí quyển. Chu trình dinh dưỡng bao gồm quá trình phân hủy xác chết; vi khuẩn chịu trách nhiệm cho giai đoạn thối rữa trong quá trình này. Trong các cộng đồng sinh vật xung quanh miệng phun thủy nhiệt và thấm lạnh, vi khuẩn ưa nhiệt cung cấp các chất dinh dưỡng cần thiết để duy trì sự sống bằng cách chuyển hóa các hợp chất hòa tan, chẳng hạn như hydro sunfua và metan, thành năng lượng. Vi khuẩn cũng sống trong các mối quan hệ cộng sinh (symbiotic) và kí sinh (parasitic) với thực vật và động vật. Hầu hết vi khuẩn chưa được đặc trưng và có nhiều loài không thể nuôi cấy trong phòng thí nghiệm. Nghiên cứu về vi khuẩn được gọi là vi khuẩn học (bacteriology), một nhánh của vi sinh vật học (microbiology).
Con người và hầu hết các loài động vật khác mang theo hàng triệu vi khuẩn. Hầu hết nằm trong ruột, và có rất nhiều trên da. Hầu hết các vi khuẩn trong và trên cơ thể là vô hại hoặc được tạo ra bởi các tác dụng bảo vệ của hệ thống miễn dịch (immune system), mặc dù nhiều vi khuẩn có lợi, đặc biệt là những vi khuẩn trong ruột. Tuy nhiên, một số loài vi khuẩn có khả năng gây bệnh và gây ra các bệnh truyền nhiễm, bao gồm bệnh tả, bệnh giang mai, bệnh than, bệnh phong, bệnh lao, bệnh uốn ván và bệnh dịch hạch. Các bệnh do vi khuẩn gây tử vong phổ biến nhất là nhiễm trùng đường hô hấp. Thuốc kháng sinh (Antibiotics) được sử dụng để điều trị các bệnh nhiễm trùng do vi khuẩn và cũng được sử dụng trong nông nghiệp, làm cho tình trạng kháng thuốc kháng sinh ngày càng trở nên nghiêm trọng. Vi khuẩn rất quan trọng trong xử lý nước thải và sự cố tràn dầu, sản xuất pho mát và sữa chua thông qua quá trình lên men, thu hồi vàng, paladi, đồng và các kim loại khác trong lĩnh vực khai thác mỏ, cũng như trong công nghệ sinh học, sản xuất thuốc kháng sinh và hóa chất khác.
Từng được coi là thực vật cấu thành lớp nấm phân hạch (schizomycetes), vi khuẩn ngày nay được xếp vào nhóm sinh vật nhân sơ. Không giống như tế bào của động vật và các sinh vật nhân thực (eukaryotes), tế bào vi khuẩn không chứa nhân và hiếm khi có các bào quan có màng bao bọc. Mặc dù thuật ngữ vi khuẩn theo truyền thống bao gồm tất cả các sinh vật nhân sơ, cách phân loại khoa học đã thay đổi sau khi phát hiện vào những năm 1990 rằng sinh vật nhân sơ bao gồm hai nhóm sinh vật rất khác nhau tiến hóa từ một tổ tiên chung cổ đại. Các lĩnh vực tiến hóa này được gọi là vi khuẩn và cổ khuẩn (archaea).
Tổ tiên của vi khuẩn là vi sinh vật đơn bào (unicellular microorganisms), là dạng sống đầu tiên xuất hiện trên Trái đất, khoảng 4 tỷ năm trước. Trong khoảng 3 tỷ năm, hầu hết các sinh vật đều ở dạng cực nhỏ, vi khuẩn và cổ khuẩn là những dạng sống thống trị. Mặc dù có tồn tại các hóa thạch vi khuẩn, chẳng hạn như stromatolite, nhưng việc thiếu hình thái đặc biệt của chúng khiến chúng không được sử dụng để kiểm tra lịch sử tiến hóa của vi khuẩn hoặc xác định thời gian xuất hiện của một loài vi khuẩn cụ thể. Tuy nhiên, trình tự gen có thể được sử dụng để tái tạo lại hệ phát sinh loài vi khuẩn (bacterial phylogeny), và những nghiên cứu này chỉ ra rằng vi khuẩn khác biệt đầu tiên khỏi dòng dõi cổ khuẩn hay sinh vật nhân thực. Tổ tiên chung gần đây nhất của vi khuẩn và cổ khuẩn có lẽ là loài siêu ưa nhiệt sống cách đây khoảng 2,5 tỷ - 3,2 tỷ năm. Sự sống sớm nhất trên cạn có thể là vi khuẩn cách đây khoảng 3,22 tỷ năm.
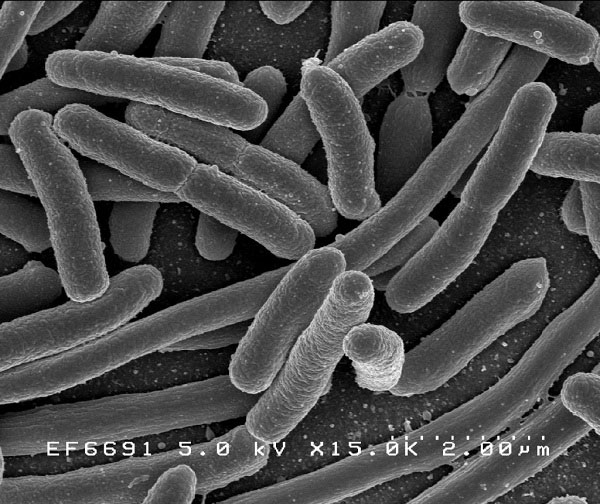
Nguồn gốc và sự tiến hóa ban đầu của vi khuẩn
Vi khuẩn cũng tham gia vào sự phân kỳ tiến hóa lớn thứ hai, đó là của cổ khuẩn và sinh vật nhân thực. Ở đây, sinh vật nhân thực là kết quả của sự xâm nhập của cổ khuẩn đại vào các liên kết nội cộng sinh với tổ tiên của các tế bào nhân thực, mà bản thân chúng có thể có liên quan đến Archaea. Điều này liên quan đến việc các tế bào tiền nhân thực hấp thụ các tế bào cộng sinh alphaproteobacteria để tạo thành ty thể (mitochondria) hoặc hydrogenosomes, vẫn được tìm thấy trong tất cả các sinh vật nhân thực đã biết ví dụ như ở động vật nguyên sinh "amitochondrial" cổ đại. Sau đó, một số sinh vật nhân thực đã chứa ty thể cũng biến đổi vi khuẩn lam như sinh vật, dẫn đến sự hình thành lục lạp (chloroplasts) ở tảo và thực vật. Đây được gọi là nhiễm trùng nội sinh nguyên phát (primary endosymbiosis).
Vi khuẩn có mặt khắp nơi, sống trong mọi môi trường sống có thể có trên hành tinh bao gồm đất, dưới nước, sâu trong vỏ Trái đất và thậm chí cả những môi trường khắc nghiệt như suối nước nóng có tính axit và chất thải phóng xạ. Có khoảng 2 × 1030 vi khuẩn trên Trái đất, tạo thành một sinh khối (biomass) chỉ xếp sau giới thực vật. Chúng có nhiều trong các hồ và đại dương, trong băng ở Bắc Cực, và các suối địa nhiệt, nơi chúng cung cấp các chất dinh dưỡng cần thiết để duy trì sự sống bằng cách chuyển đổi các hợp chất hòa tan, chẳng hạn như hydro sunfua và metan thành năng lượng.

Môi trường sống của vi khuẩn
Vi khuẩn sống trên cả thực vật và động vật. Hầu hết không gây bệnh, có lợi cho môi trường sống và cần thiết cho sự sống. Đất là nguồn vi khuẩn phong phú và một vài gam chứa khoảng một nghìn triệu vi khuẩn trong số đó. Tất cả chúng đều rất cần thiết cho hệ sinh thái đất, phân hủy chất thải độc hại và tái chế chất dinh dưỡng. Chúng thậm chí còn được tìm thấy trong khí quyển và một mét khối không khí chứa khoảng một trăm triệu tế bào vi khuẩn. Các đại dương và biển có khoảng 3 x 1026 vi khuẩn cung cấp tới 50% lượng oxy mà con người hít thở. Chỉ có khoảng 2% các loài vi khuẩn đã được nghiên cứu đầy đủ.
Vi khuẩn ưa nhiệt
|
Môi trường sống |
Giống loài |
|
Lạnh (âm 15 ° C Nam Cực) |
Cryptoendoliths |
|
Nóng (mạch nước phun 70–100 ° C) |
Thermus aquus |
|
Bức xạ, 5M Rad |
Deinococcus radiodurans |
|
Muối, 47% muối (Biển Chết, Hồ Muối Lớn) |
một số loài |
|
Axit pH 3 |
một số loài |
|
Độ pH kiềm 12,8 |
vi khuẩn betaproteobacteria |
|
Không gian (6 năm trên vệ tinh của NASA) |
Bacillus subtilis |
|
3,2 km dưới lòng đất |
một số loài |
Vi khuẩn hiển thị rất nhiều hình dạng và kích thước. Tế bào vi khuẩn có kích thước bằng 1/10 tế bào nhân thực và thường có chiều dài 0,5–5,0 micromet. Tuy nhiên, một số loài có thể nhìn thấy bằng mắt thường. Ví dụ: Thiomargarita namibiensis dài tới nửa milimet, Epulopiscium fishelsoni đạt 0,7 mm và Thiomargarita phóng đại có thể dài tới 2 cm. Trong số các vi khuẩn nhỏ nhất là các thành viên của chi Mycoplasma, chỉ có kích thước 0,3 micromet nhỏ bằng các vi rút (virus) lớn nhất. Một số vi khuẩn thậm chí có thể nhỏ hơn, nhưng những vi khuẩn siêu nhỏ này chưa được nghiên cứu kỹ lưỡng.
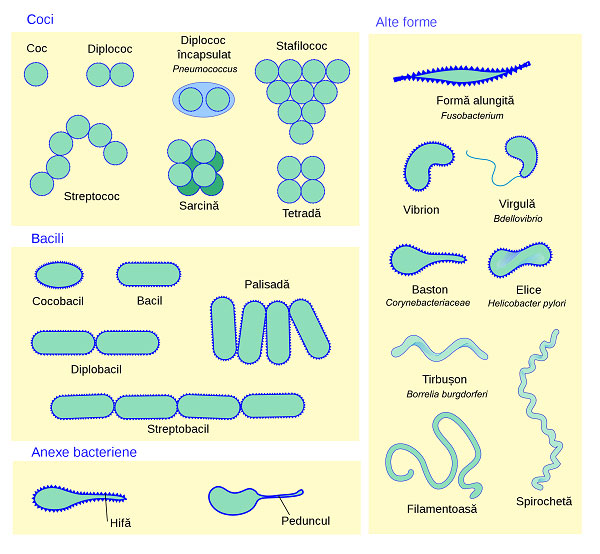
Cấu tạo hình thái học của vi khuẩn
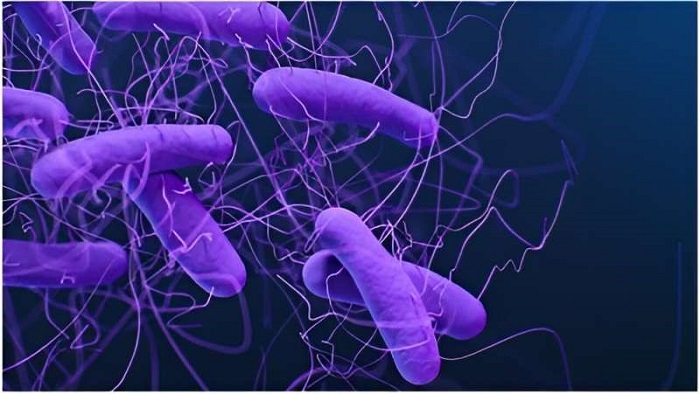
Hầu hết các loài vi khuẩn đều có hình cầu, được gọi là cầu khuẩn (cocci, từ tiếng Hy Lạp “kókkos”) hoặc hình que, được gọi là trực khuẩn (bacilli, từ tiếng Latinh là “baculus”). Một số vi khuẩn được gọi là vibrio, có hình dạng giống như hình que hơi cong hoặc hình dấu phẩy. Những xoắn khuẩn khác có thể có hình xoắn ốc được gọi là spirilla hoặc cuộn chặt được gọi là spirochaetes. Một số ít các hình dạng bất thường khác đã được mô tả, chẳng hạn như vi khuẩn hình ngôi sao. Sự đa dạng về hình dạng này được xác định bởi vi khuẩn thành tế bào (cell wall) và bộ xương tế bào (cytoskeleton). Nó rất quan trọng vì có thể ảnh hưởng đến khả năng vi khuẩn thu nhận chất dinh dưỡng, bám vào bề mặt, bơi qua chất lỏng và thoát khỏi động vật ăn thịt.
Hầu hết các loài vi khuẩn tồn tại đơn giản dưới dạng tế bào đơn lẻ; những người khác liên kết theo các kiểu đặc trưng: Vi khuẩn lậu cầu tạo thành lưỡng bội (cặp), liên cầu tạo thành chuỗi và tụ cầu nhóm với nhau trong các cụm hình chùm nho. Vi khuẩn cũng có thể nhóm lại để tạo thành các cấu trúc đa bào lớn hơn, chẳng hạn như các sợi dài của các loài Actinomycetota, các tập hợp của các loài Myxobacteria và các sợi nấm phức tạp của các loài Streptomyces. Những cấu trúc đa bào này thường chỉ được nhìn thấy trong những điều kiện nhất định.
Ví dụ: khi thiếu axit amin, vi khuẩn myxobacteria phát hiện ra các tế bào xung quanh trong một quá trình được gọi là cảm ứng số lượng (quorum sensing), di chuyển về phía nhau và tập hợp lại để tạo thành quả thể (fruiting bodies) dài tới 500 micromet và chứa khoảng 100.000 tế bào vi khuẩn. Trong các quả thể này, vi khuẩn thực hiện các nhiệm vụ riêng biệt. Ví dụ: khoảng 1/10 tế bào di chuyển đến đỉnh của quả thể và biệt hóa thành trạng thái ngủ đông chuyên biệt được gọi là myxospore, có khả năng chống lại sự khô và các điều kiện môi trường bất lợi khác cao hơn.
Vi khuẩn thường bám vào các bề mặt và tạo thành các tập hợp dày đặc được gọi là màng sinh học (biofilms) và các thành tạo lớn hơn được gọi là thảm vi sinh vật (microbial mats). Những màng sinh học và thảm này có thể dày từ vài micromet đến độ sâu nửa mét và có thể chứa nhiều loài vi khuẩn, sinh vật nguyên sinh (protists) và cổ khuẩn.
Vi khuẩn sống trong màng sinh học thể hiện sự sắp xếp phức tạp của các tế bào và các thành phần ngoại bào, tạo thành các cấu trúc thứ cấp, qua đó có mạng lưới các kênh để cho phép chất dinh dưỡng khuếch tán tốt hơn. Trong môi trường tự nhiên, chẳng hạn như đất hoặc bề mặt của thực vật, phần lớn vi khuẩn bám vào các bề mặt trong màng sinh học.
Màng sinh học cũng rất quan trọng trong y học, vì những cấu trúc này thường xuất hiện trong các trường hợp nhiễm vi khuẩn mãn tính hoặc nhiễm trùng các thiết bị y tế được cấy ghép và vi khuẩn được bảo vệ trong màng sinh học khó bị tiêu diệt hơn nhiều so với vi khuẩn được phân lập riêng lẻ.
Tế bào vi khuẩn được bao bọc bởi màng tế bào (cell membrane), màng này được tạo ra chủ yếu từ các phospholipid. Màng này bao bọc các chất bên trong tế bào và hoạt động như một hàng rào để giữ các chất dinh dưỡng, protein và các thành phần thiết yếu khác của tế bào chất bên trong tế bào. Không giống như tế bào nhân thực, vi khuẩn thường thiếu các cấu trúc liên kết màng lớn trong chất tế bào của chúng như nhân, ty thể, lục lạp và các bào quan khác có trong tế bào nhân thực. Tuy nhiên, một số vi khuẩn có các bào quan liên kết với protein trong tế bào chất của vi khuẩn giúp ngăn các khía cạnh trao đổi chất của vi khuẩn, chẳng hạn như carboxysome. Ngoài ra, vi khuẩn có một bộ xương tế bào đa thành phần để kiểm soát sự bản địa hóa của các protein và axit nucleic trong tế bào và quản lý quá trình phân chia tế bào (cell division).
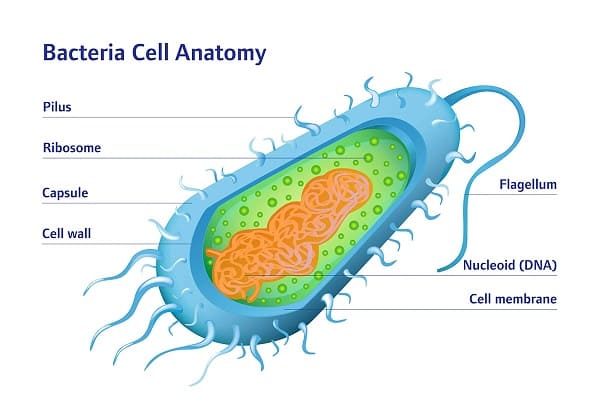
Cấu trúc tế bào của vi khuẩn
Nhiều phản ứng sinh hóa xảy ra do thang độ nồng độ (concentration gradients) qua các màng tạo ra năng lượng tương tự như pin. Sự thiếu hụt màng bên trong nói chung ở vi khuẩn là những phản ứng vận chuyển electron xảy ra qua màng tế bào, giữa tế bào chất (cytoplasm) và bên ngoài tế bào hoặc giữa các ngoại chất (periplasm). Tuy nhiên, ở nhiều vi khuẩn quang hợp, màng sinh chất (plasma membrane) rất gấp và lấp đầy phần lớn tế bào bằng các lớp màng thu nhận ánh sáng. Những phức hợp thu nhận ánh sáng này thậm chí có thể hình thành cấu trúc bao bọc lipid được gọi là lục lạp trong vi khuẩn lưu huỳnh xanh. Màng sinh chất của tế bào vi khuẩn không có Colesteron.
Vi khuẩn không có nhân bao quanh màng, và vật chất di truyền của chúng thường là một nhiễm sắc thể (transduction) hình tròn đơn của DNA nằm trong tế bào chất trong một cơ thể có hình dạng bất thường được gọi là vùng nhân (nucleoid). Vùng nhân chứa nhiễm sắc thể với các protein và RNA liên kết của nó. Giống như tất cả các sinh vật khác, vi khuẩn chứa ribosome để sản xuất protein, nhưng cấu trúc của ribosome vi khuẩn khác với cấu trúc của sinh vật nhân thực và cổ khuẩn.
Một số vi khuẩn tạo ra các hạt lưu trữ chất dinh dưỡng nội bào (intracellular), chẳng hạn như glycogen, polyphosphat, lưu huỳnh hoặc polyhydroxyalkanoat. Các vi khuẩn như vi khuẩn lam quang hợp, tạo ra các không bào khí (gas vacuoles) bên trong, chúng sử dụng để điều chỉnh sự nổi của chúng, cho phép chúng di chuyển lên hoặc xuống các lớp nước với cường độ ánh sáng và mức độ dinh dưỡng khác nhau.
Xung quanh bên ngoài của màng tế bào là thành tế bào. Thành tế bào vi khuẩn cấu tạo chủ yếu từ peptidoglycan (còn gọi là murein), được tạo ra từ các chuỗi polysaccharid liên kết chéo với nhau bởi các peptit chứa D- axit amin. Thành tế bào vi khuẩn khác với thành tế bào của thực vật và nấm, được cấu tạo từ xenlulo và kitin, tương ứng. Thành tế bào của vi khuẩn cũng khác với thành tế bào của cổ khuẩn, không chứa peptidoglycan. Chức năng của thành tế bào vi khuẩn là cần thiết cho sự tồn tại của nhiều vi khuẩn và kháng sinh penicillin (do một loại nấm có tên là Penicillium sinh ra) có khả năng tiêu diệt vi khuẩn bằng cách ức chế một bước trong quá trình tổng hợp peptidoglycan.
Nói một cách rộng ra, có hai loại thành tế bào khác nhau ở vi khuẩn, phân loại vi khuẩn thành vi khuẩn Gram dương (Gram-positive) và vi khuẩn Gram âm (Gram-negative). Tên gọi bắt nguồn từ phản ứng của tế bào với nhuộm Gram, một thử nghiệm lâu đời để phân loại các loài vi khuẩn.
Vi khuẩn gram dương có thành tế bào dày chứa nhiều lớp axit peptidoglycan và teichoic. Ngược lại, vi khuẩn Gram âm có thành tế bào tương đối mỏng bao gồm một vài lớp peptidoglycan được bao bọc bởi một màng lipid thứ hai chứa lipopolysaccharid và lipoprotein. Hầu hết các vi khuẩn có thành tế bào Gram âm, và chỉ các thành viên của nhóm Bacillota và actinomycetota (trước đây được gọi là vi khuẩn Gram dương có G + C thấp và G + C cao) có sự sắp xếp Gram dương thay thế. Những khác biệt về cấu trúc này có thể tạo ra sự khác biệt về tính nhạy cảm với kháng sinh. Ví dụ: vancomycin chỉ có thể tiêu diệt vi khuẩn Gram dương và không có hiệu quả đối với các vi khuẩn gây bệnh Gram âm, chẳng hạn như Haemophilus influenzae hoặc Pseudomonas aeruginosa. Một số vi khuẩn có cấu trúc thành tế bào không phải là Gram dương hoặc Gram âm cổ điển. Điều này bao gồm các vi khuẩn quan trọng về mặt lâm sàng như vi khuẩn mycobacteria có thành tế bào peptidoglycan dày như vi khuẩn Gram dương, nhưng cũng có lớp lipid bên ngoài thứ hai.
Ở nhiều loài vi khuẩn, một lớp S gồm các phân tử protein xếp thành từng mảng chắc chắn bao phủ bên ngoài tế bào. Lớp này cung cấp sự bảo vệ hóa học và vật lý cho bề mặt tế bào và có thể hoạt động như một hàng rào khuếch tán đại phân tử (macromolecular). Các lớp S có chức năng đa dạng và được biết là yếu tố độc lực ở các loài Campylobacter và chứa các enzym bề mặt trong Bacillus stearothermophilus.
Trùng roi (Flagella) là cấu trúc protein cứng, có đường kính khoảng 20 nanomet và chiều dài lên đến 20 micromet được sử dụng để vận động. Trùng roi được thúc đẩy bởi năng lượng được giải phóng bằng cách chuyển các ion xuống một gradient điện hóa qua màng tế bào.
Fimbriae (đôi khi được gọi là "đính kèm pili") là những sợi protein nhỏ, thường có đường kính từ 2–10 nanomet và chiều dài lên đến vài micromet. Chúng phân bố trên bề mặt tế bào và trông giống như những sợi lông mịn khi nhìn dưới kính hiển vi điện tử. Fimbriae được cho là có liên quan đến việc bám vào bề mặt rắn hoặc với các tế bào khác và rất cần thiết cho độc lực (virulence) của một số vi khuẩn gây bệnh. Pili (Pilus) là phần phụ của tế bào, lớn hơn một chút so với fimbriae, có thể chuyển vật chất di truyền giữa các tế bào vi khuẩn trong một quá trình được gọi là tiếp hợp (conjugation), nơi chúng được gọi là pili tiếp hợp hoặc pili giới tính. Chúng cũng có thể tạo ra chuyển động mà chúng được gọi là pili loại IV.
Glycocalyx được tạo ra bởi nhiều vi khuẩn để bao quanh tế bào của chúng và khác nhau về độ phức tạp về cấu trúc: từ một lớp chất nhờn không được tổ chức gồm các chất cao phân tử ngoại bào đến một viên nang có cấu trúc cao. Những cấu trúc này có thể bảo vệ các tế bào khỏi bị nhấn chìm bởi các tế bào nhân thực như đại thực bào (macrophages - một phần của hệ thống miễn dịch của con người). Chúng cũng có thể hoạt động như kháng nguyên và tham gia vào quá trình nhận dạng tế bào, cũng như hỗ trợ gắn vào các bề mặt và hình thành màng sinh học.
Cấu tạo các cấu trúc ngoại bào này phụ thuộc vào hệ thống bài tiết của vi khuẩn. Các protein này chuyển từ tế bào chất vào ngoại chất hoặc vào môi trường xung quanh tế bào. Nhiều loại hệ thống bài tiết đã được biết đến và những cấu trúc này thường rất cần thiết cho độc lực của vi khuẩn gây bệnh và đang được nghiên cứu chuyên sâu.
Một số chi vi khuẩn Gram dương, chẳng hạn như Bacillus, Clostridium, Sporohalobacter, Anaerobacter và Heliobacterium, có thể hình thành các cấu trúc ngủ đông, có khả năng kháng cao được gọi là nội bào tử (endospores). Nội bào tử phát triển trong tế bào chất của tế bào và chỉ có một nội bào tử duy nhất phát triển trong mỗi tế bào. Mỗi nội bào tử chứa một lõi DNA và ribosome được bao quanh bởi một lớp vỏ và được bảo vệ bởi một lớp áo cứng nhiều lớp bao gồm peptidoglycan và nhiều loại protein.
Các bào tử nội bào tử không cho thấy sự trao đổi chất có thể phát hiện được và có thể tồn tại những điều kiện khắc nghiệt về vật lý và hóa học, chẳng hạn như mức độ cao của tia UV, bức xạ gamma, chất tẩy rửa, chất khử trùng, nhiệt, đóng băng, áp suất và hút ẩm. Trong trạng thái không hoạt động này, những sinh vật này có thể vẫn tồn tại trong hàng triệu năm và nội bào tử thậm chí còn cho phép vi khuẩn sống sót khi tiếp xúc với chân không và bức xạ trong không gian, có thể vi khuẩn có thể phân bố khắp Vũ trụ bởi bụi không gian, thiên thạch, tiểu hành tinh, sao chổi, hành tinh hoặc thông qua panspermia định hướng. Vi khuẩn hình thành nội bào tử cũng có thể gây bệnh: ví dụ, bệnh than có thể lây nhiễm khi hít phải nội bào tử Bacillus anthracis và nhiễm nội bào tử Clostridium tetani gây ra bệnh uốn ván, giống như bệnh ngộ độc gây ra độc tố do vi khuẩn phát triển từ bào tử tiết ra. Nhiễm trùng Clostridioides difficile là một vấn đề trong cơ sở chăm sóc sức khỏe cũng do vi khuẩn hình thành bào tử gây ra.
Vi khuẩn thể hiện rất nhiều kiểu trao đổi chất (metabolism). Sự phân bố các đặc điểm trao đổi chất trong một nhóm vi khuẩn theo truyền thống được sử dụng để xác định phân loại của chúng, nhưng những đặc điểm này thường không tương ứng với các phân loại di truyền hiện đại. Sự trao đổi chất của vi khuẩn được phân loại thành các nhóm dinh dưỡng trên cơ sở ba tiêu chí chính: nguồn năng lượng, các chất cho electron được sử dụng và nguồn cacbon được sử dụng để tăng trưởng.
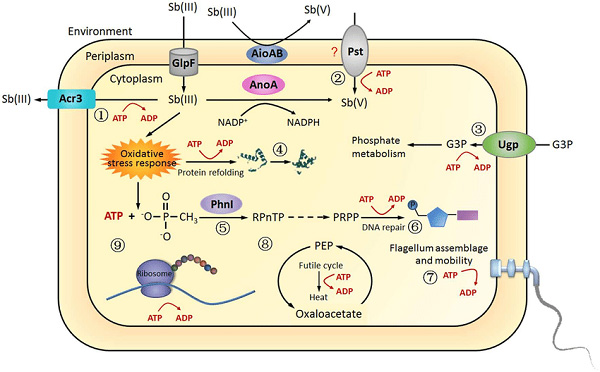
Sự trao đổi chất (Metabolism)
Vi khuẩn hoặc lấy năng lượng từ ánh sáng bằng cách sử dụng quang hợp (photosynthesis) quá trình này được gọi là quang dưỡng (phototrophy). Hoặc bằng cách phá vỡ các hợp chất hóa học bằng cách sử dụng quá trình oxy hóa (oxidation) quá trình này được gọi là quang hóa (chemotrophy). Nhiều vi khuẩn sử dụng các hợp chất hóa học làm nguồn năng lượng bằng cách chuyển các electron từ chất cho electron nhất định đến chất nhận electron cuối cùng trong phản ứng oxy hóa khử. Phản ứng này giải phóng năng lượng có thể được sử dụng để thúc đẩy quá trình trao đổi chất. Các sinh vật hóa học được phân chia nhiều hơn theo các loại hợp chất mà chúng sử dụng để chuyển electron. Vi khuẩn sử dụng các hợp chất vô cơ như hydro, carbon monoxide hoặc amoniac làm nguồn electron được gọi là sinh vật tự dưỡng (lithotrophs), trong khi những sinh vật sử dụng các hợp chất hữu cơ được gọi là sinh vật hữu cơ (organotrophs). Các hợp chất được sử dụng để nhận electron cũng được sử dụng để phân loại vi khuẩn: sinh vật hiếu khí sử dụng oxy làm chất nhận electron cuối cùng, trong khi sinh vật kỵ khí sử dụng các hợp chất khác như nitrat, sunfat hoặc carbon dioxide.
Nhiều vi khuẩn lấy carbon của chúng từ carbon hữu cơ khác, được gọi là dị dưỡng (heterotrophy). Những loài khác như vi khuẩn lam và một số vi khuẩn màu tía là tự dưỡng, có nghĩa là chúng thu được carbon tế bào bằng cách cố định carbon dioxide. Trong những trường hợp bất thường, khí metan có thể được vi khuẩn dị dưỡng sử dụng làm nguồn cung cấp electron và chất nền cho quá trình đồng hóa cacbon.
Các loại dinh dưỡng trong quá trình trao đổi chất của vi khuẩn:
|
Loại dinh dưỡng |
Nguồn năng lượng |
Nguồn carbon |
Các ví dụ |
|
Sinh vật quang dưỡng |
Ánh sáng mặt trời |
Các hợp chất hữu cơ (quang dưỡng) hoặc cố định cacbon (quang tự dưỡng) |
Vi khuẩn lam, vi khuẩn lưu huỳnh màu xanh lục, vi khuẩn Chloroflexota hoặc vi khuẩn màu tím |
|
Lithotrophs |
Hợp chất vô cơ |
Các hợp chất hữu cơ (sinh vật dị dưỡng) hoặc cố định cacbon (sinh vật tự dưỡng) |
Thermodesulfobacteriota, Hydrogenophilaceae hoặc Nitrospirae |
|
Organotrophs |
Hợp chất hữu cơ |
Các hợp chất hữu cơ (chemoheterotrophs) hoặc cố định carbon (chemoautotrophs) |
Bacillus, Clostridium, hoặc Enterobacteriaceae |
Theo nhiều cách, sự trao đổi chất của vi khuẩn cung cấp những đặc điểm có ích cho sự ổn định sinh thái và cho xã hội loài người. Một ví dụ là một số vi khuẩn được gọi là diazotrophs có khả năng cố định khí nitơ bằng cách sử dụng enzyme nitrogenase. Đặc điểm quan trọng với môi trường này có thể được tìm thấy ở vi khuẩn thuộc hầu hết các loại trao đổi chất được liệt kê ở trên. Điều này dẫn đến các quá trình quan trọng về mặt sinh thái của quá trình khử nitơ, khử sunfat và tạo acetogenes, tương ứng. Quá trình trao đổi chất của vi khuẩn cũng rất quan trọng trong các phản ứng sinh học đối với ô nhiễm; ví dụ như vi khuẩn khử sunfat chịu trách nhiệm phần lớn trong việc tạo ra các dạng thủy ngân có độc tính cao (metyl và dimethylmercury) trong môi trường. Vi khuẩn kỵ khí không hô hấp sử dụng quá trình lên men để tạo ra năng lượng và giảm năng lượng, tiết ra các sản phẩm phụ trao đổi chất (chẳng hạn như ethanol trong sản xuất bia) làm chất thải. Các vi khuẩn kỵ khí nuôi cấy có thể chuyển đổi giữa quá trình lên men và các chất nhận electron đầu cuối khác nhau tùy thuộc vào điều kiện môi trường mà chúng tự tìm thấy.
Không giống như ở sinh vật đa bào (multicellular organisms), sự gia tăng kích thước tế bào (tăng trưởng tế bào) và sinh sản bằng cách phân chia tế bào được liên kết chặt chẽ trong sinh vật đơn bào (unicellular organisms). Vi khuẩn phát triển đến một kích thước cố định và sau đó sinh sản thông qua phân hạch nhị phân (binary fission), là một hình thức sinh sản vô tính (asexual reproduction). Trong điều kiện tối ưu, vi khuẩn có thể phát triển và phân chia cực kỳ nhanh chóng, một số quần thể vi khuẩn có thể nhân đôi nhanh nhất cứ sau 17 phút. Trong quá trình phân chia tế bào, hai bản sao giống hệt nhau tế bào con được tạo ra. Một số vi khuẩn, trong khi vẫn sinh sản vô tính, hình thành các cấu trúc sinh sản phức tạp hơn giúp phân tán các tế bào con mới hình thành. Các ví dụ bao gồm sự hình thành quả thể do vi khuẩn myxobacteria và sự hình thành sợi nấm trên không của các loài Streptomyces, hoặc nảy chồi. Quá trình nảy chồi liên quan đến việc một tế bào hình thành một phần lồi bị vỡ ra và tạo ra một tế bào con.
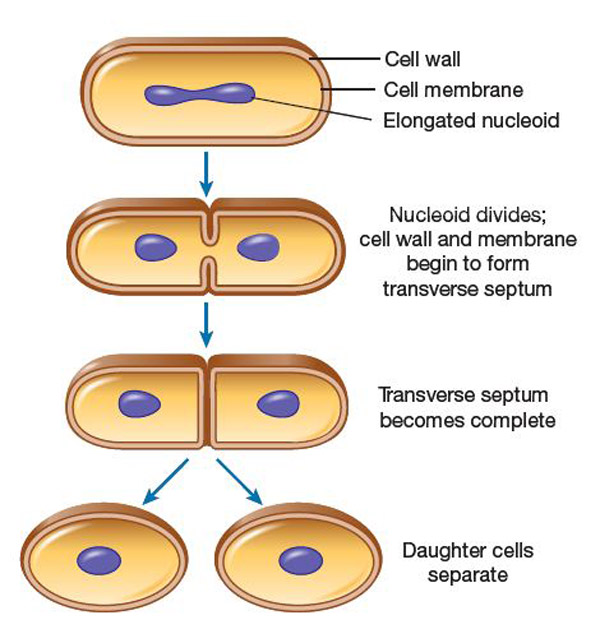
Tăng trưởng và sinh sản của vi khuẩn
Trong phòng thí nghiệm, vi khuẩn thường được nuôi cấy bằng môi trường rắn hoặc lỏng. Môi trường nuôi cấy rắn, chẳng hạn như đĩa thạch, được sử dụng để phân lập các dòng vi khuẩn nuôi cấy thuần túy. Tuy nhiên, môi trường tăng trưởng dạng lỏng được sử dụng khi cần đo sự phát triển hoặc khối lượng lớn tế bào. Sự phát triển trong môi trường lỏng được khuấy trộn xảy ra dưới dạng huyền phù tế bào (cell suspension) đồng đều, làm cho các mẫu cấy dễ dàng phân chia và chuyển giao, mặc dù việc phân lập vi khuẩn đơn lẻ khỏi môi trường lỏng là rất khó. Việc sử dụng môi trường chọn lọc (môi trường có bổ sung hoặc thiếu chất dinh dưỡng cụ thể, hoặc có thêm kháng sinh) có thể giúp xác định các sinh vật cụ thể.
Hầu hết các kỹ thuật nuôi cấy vi khuẩn trong phòng thí nghiệm đều sử dụng hàm lượng chất dinh dưỡng cao để tạo ra một lượng lớn tế bào với giá rẻ và nhanh chóng. Tuy nhiên, trong môi trường tự nhiên, chất dinh dưỡng bị hạn chế, có nghĩa là vi khuẩn không thể tiếp tục sinh sản vô thời hạn. Sự hạn chế về chất dinh dưỡng này đã dẫn đến sự phát triển của các chiến lược tăng trưởng khác nhau (lý thuyết chọn lọc r/K). Một số sinh vật có thể phát triển cực kỳ nhanh chóng khi các chất dinh dưỡng có sẵn, chẳng hạn như sự hình thành của tảo (vi khuẩn lam) nở hoa thường xảy ra trong các hồ vào mùa hè. Các sinh vật khác có khả năng thích nghi với môi trường khắc nghiệt, chẳng hạn như sản xuất nhiều loại thuốc kháng sinh bởi streptomyces ức chế sự phát triển của vi sinh vật cạnh tranh. Trong tự nhiên, nhiều sinh vật sống trong các cộng đồng (ví dụ: màng sinh học) có thể cho phép tăng cường cung cấp chất dinh dưỡng và bảo vệ khỏi các áp lực môi trường. Những mối quan hệ này có thể cần thiết cho sự phát triển của một sinh vật hoặc nhóm sinh vật cụ thể (sinh vật tổng hợp).
Sự phát triển của vi khuẩn theo bốn giai đoạn. Khi một quần thể vi khuẩn lần đầu tiên bước vào một môi trường có chất dinh dưỡng cao cho phép phát triển, các tế bào cần phải thích nghi với môi trường mới của chúng.
- Giai đoạn đầu của sinh trưởng là giai đoạn trễ (lag phase), giai đoạn phát triển chậm lại khi các tế bào đang thích nghi với môi trường dinh dưỡng cao và chuẩn bị cho sự phát triển nhanh. Giai đoạn trễ có tốc độ sinh tổng hợp cao, vì các protein cần thiết cho sự phát triển nhanh chóng được sản xuất.
- Giai đoạn tăng trưởng thứ hai là giai đoạn logarit (logarithmic phase), còn được gọi là giai đoạn hàm mũ. Giai đoạn log được đánh dấu bằng tốc độ tăng trưởng nhanh chóng theo cấp số nhân. Tốc độ tế bào phát triển trong giai đoạn này được gọi là tốc độ tăng trưởng (k), và thời gian để các ô nhân đôi được gọi là thời gian tạo (g). Trong giai đoạn log, các chất dinh dưỡng được chuyển hóa ở tốc độ tối đa cho đến khi một trong các chất dinh dưỡng cạn kiệt và bắt đầu hạn chế sự tăng trưởng.
- Giai đoạn thứ ba của sự tăng trưởng là giai đoạn tĩnh (stationary phase) là do các chất dinh dưỡng cạn kiệt. Các tế bào giảm hoạt động trao đổi chất và tiêu thụ các protein không thiết yếu của tế bào. Giai đoạn tĩnh là sự chuyển đổi từ trạng thái tăng trưởng nhanh sang trạng thái phản ứng căng thẳng và có sự gia tăng biểu hiện của các gen liên quan đến quá trình sửa chữa DNA, chuyển hóa chất chống oxy hóa và vận chuyển chất dinh dưỡng.
- Giai đoạn cuối cùng là giai đoạn chết (death phase), là giai đoạn vi khuẩn cạn kiệt chất dinh dưỡng và chết.
Hầu hết các vi khuẩn có một nhiễm sắc thể hình tròn duy nhất có thể có kích thước chỉ từ 160.000 cặp bazơ ở vi khuẩn nội cộng sinh Carsonella ruddii, đến 12.200.000 cặp bazơ (12,2 Mbp) ở vi khuẩn sống trong đất Sorangium cellulosum. Có nhiều ngoại lệ cho điều này, ví dụ một số loài Streptomyces và Borrelia chứa một nhiễm sắc thể tuyến tính đơn, trong khi một số loài Vibrio chứa nhiều hơn một nhiễm sắc thể. Vi khuẩn cũng có thể chứa plasmid, các phân tử DNA ngoài nhiễm sắc thể nhỏ có thể chứa các gen cho các chức năng hữu ích khác nhau như khả năng kháng kháng sinh, khả năng trao đổi chất hoặc các yếu tố độc lực khác nhau.
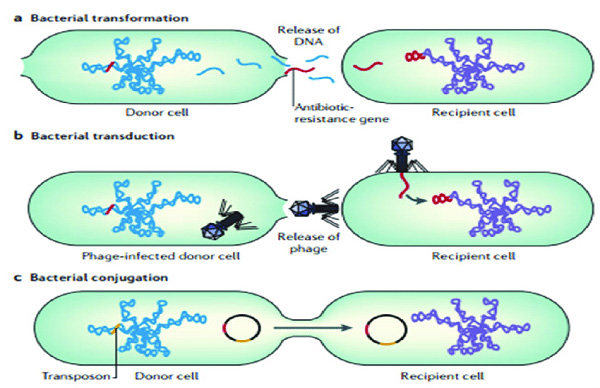
Di truyền học của vi khuẩn (Genetics)
Bộ gen của vi khuẩn thường mã hóa từ vài trăm đến vài nghìn gen. Các gen trong bộ gen của vi khuẩn thường là một đoạn DNA liên tục đơn lẻ và mặc dù có một số loại intron khác nhau tồn tại ở vi khuẩn, chúng hiếm hơn nhiều so với ở sinh vật nhân thực.
Vi khuẩn là sinh vật vô tính, thừa kế một bản sao giống hệt bộ gen của bố mẹ. Tuy nhiên, tất cả các vi khuẩn có thể tiến hóa bằng cách chọn lọc những thay đổi đối với DNA vật chất di truyền của chúng do tái tổ hợp hoặc đột biến gen (mutations). Đột biến đến từ các lỗi được thực hiện trong quá trình sao chép DNA hoặc do tiếp xúc với các đột biến. Tỷ lệ đột biến rất khác nhau giữa các loài vi khuẩn khác nhau và thậm chí giữa các dòng vô tính khác nhau của một loài vi khuẩn. Những thay đổi di truyền trong bộ gen của vi khuẩn đến từ đột biến ngẫu nhiên trong quá trình sao chép hoặc "đột biến hướng căng thẳng" (stress-directed mutation), trong đó các gen liên quan đến một quá trình hạn chế tăng trưởng cụ thể có tỷ lệ đột biến tăng lên.
Một số vi khuẩn cũng chuyển vật chất di truyền giữa các tế bào. Điều này có thể xảy ra theo ba cách chính:
- Đầu tiên, vi khuẩn có thể lấy DNA ngoại sinh từ môi trường của chúng, trong một quá trình được gọi là biến nạp (transformation). Nhiều vi khuẩn có thể tiếp nhận DNA từ môi trường một cách tự nhiên, trong khi những vi khuẩn khác phải được thay đổi về mặt hóa học để khiến chúng tiếp nhận DNA. Sự phát triển năng lực trong tự nhiên thường gắn liền với các điều kiện môi trường khắc nghiệt, và dường như là một sự thích nghi để tạo điều kiện sửa chữa các tổn thương DNA trong tế bào nhận.
- Thứ hai, vi khuẩn chuyển vật chất di truyền là bằng cách tải nạp (transduction), khi tích hợp thực khuẩn đưa DNA ngoại lai vào nhiễm sắc thể. Nhiều loại xạ khuẩn tồn tại, một số chỉ lây nhiễm và ngăn chặn vi khuẩn chủ của chúng, trong khi những loại khác chèn vào nhiễm sắc thể của vi khuẩn. Vi khuẩn chống lại sự xâm nhiễm của phage thông qua hệ thống sửa đổi giới hạn làm suy giảm DNA ngoại lai và một hệ thống sử dụng trình tự CRISPR để giữ lại các đoạn gen của phage mà vi khuẩn đã tiếp xúc trong quá khứ, điều này cho phép chúng ngăn chặn sự nhân lên của virus thông qua một hình thức can thiệp RNA.
- Phương pháp chuyển gen thứ ba là tiếp hợp, nhờ đó DNA được chuyển qua tiếp xúc trực tiếp với tế bào. Trong các trường hợp thông thường, quá trình tải nạp, tiếp hợp và biến nạp liên quan đến việc chuyển DNA giữa các vi khuẩn riêng lẻ của cùng một loài, nhưng đôi khi sự chuyển giao có thể xảy ra giữa các cá thể thuộc các loài vi khuẩn khác nhau và điều này có thể gây ra những hậu quả đáng kể, chẳng hạn như chuyển giao kháng thuốc kháng sinh. Trong những trường hợp như vậy, việc thu nhận gen từ vi khuẩn khác hoặc từ môi trường được gọi là chuyển gen ngang (horizontal gene transfer) và có thể phổ biến trong điều kiện tự nhiên.
Nhiều vi khuẩn có tính di động (motile), có thể tự di chuyển bằng nhiều cơ chế khác nhau. Nghiên cứu cho thấy di chuyển tốt nhất trong số này là hình roi (flagella), các sợi dài được quay bởi một bộ phần ở đuôi tạo ra chuyển động giống như cánh quạt. Trùng roi (flagellum) được tạo ra từ khoảng 20 protein, với khoảng 30 protein khác cần thiết để điều hòa và lắp ráp. Trùng roi là một cấu trúc quay được điều khiển bởi một bộ phận có thể đảo ngược ở đế sử dụng gradien điện hóa qua màng để cung cấp năng lượng.

Sự chuyển động của vi khuẩn (Movement)
Vi khuẩn có thể sử dụng trùng roi theo nhiều cách khác nhau để tạo ra các kiểu di chuyển khác nhau. Nhiều vi khuẩn (chẳng hạn như E. coli) có hai hình thức di chuyển riêng biệt: di chuyển về phía trước (bơi lội) và nhào lộn. Cú lộn nhào cho phép họ định hướng lại và làm cho chuyển động của họ trở thành một bước đi ngẫu nhiên ba chiều. Các loài vi khuẩn khác nhau về số lượng và sự sắp xếp của các roi trên bề mặt của chúng; một số có một trùng roi đơn (monotrichous), một trùng roi ở mỗi đầu (amphitrichous), các cụm roi ở các cực của tế bào (lophotrichous), trong khi những người khác có các roi phân bố trên toàn bộ bề mặt của tế bào phúc mạc (peritrichous). Trùng roi của một nhóm vi khuẩn độc nhất, xoắn khuẩn, được tìm thấy giữa hai màng trong không gian ngoại chất. Chúng có một cơ thể xoắn đặc biệt xoay quanh khi nó di chuyển.
Hai loại chuyển động khác của vi khuẩn được gọi là chuyển động co giật (twitching motility) dựa vào cấu trúc được gọi là tiêm mao loại IV và chuyển động lượn sử dụng các cơ chế khác. Khi chuyển động co giật, tiêm mao giống hình que vươn ra khỏi tế bào, liên kết với một số chất nền, sau đó co lại, kéo tế bào về phía trước.
Vi khuẩn di động bị thu hút hoặc đẩy lùi bởi một số kích thích trong các hành vi được gọi là “taxes”: chúng bao gồm điều hòa hóa học, điều hòa quang học, taxi năng lượng và điều khiển từ tính. Trong một nhóm đặc biệt, vi khuẩn myxobacteria, các vi khuẩn riêng lẻ di chuyển cùng nhau để tạo thành các đợt tế bào sau đó phân hóa để hình thành quả thể chứa bào tử. Vi khuẩn myxobacteria chỉ di chuyển khi ở trên bề mặt rắn, không giống như E. coli, di chuyển trong môi trường lỏng hoặc rắn.
Một số loài Listeria và Shigella di chuyển bên trong tế bào vật chủ bằng cách chiếm đoạt bộ xương tế bào, bộ xương thường được sử dụng để di chuyển các bào quan bên trong tế bào. Bằng cách thúc đẩy quá trình polyme hóa actin ở một cực của tế bào, chúng có thể hình thành một loại đuôi đẩy chúng qua tế bào chất của tế bào chủ.
Một số ít vi khuẩn có hệ thống hóa học tạo ra ánh sáng. Hiện tượng phát quang sinh học này thường xảy ra ở vi khuẩn sống chung với cá và ánh sáng có thể dùng để thu hút cá hoặc các động vật lớn khác.
Vi khuẩn thường hoạt động như một tập hợp đa bào được gọi là màng sinh học, trao đổi nhiều loại tín hiệu phân tử để liên lạc giữa các tế bào và tham gia vào các hành vi đa bào phối hợp.
Các lợi ích chung của sự hợp tác đa bào bao gồm sự phân chia lao động của tế bào, tiếp cận các nguồn tài nguyên mà các tế bào đơn lẻ không thể sử dụng hiệu quả, bảo vệ tập thể chống lại các chất đối kháng và tối ưu hóa sự tồn tại của quần thể bằng cách phân hóa thành các loại tế bào riêng biệt. Ví dụ: vi khuẩn trong màng sinh học có thể có khả năng kháng các tác nhân kháng khuẩn tăng hơn 500 lần so với vi khuẩn "phiêu sinh" (planktonic) riêng lẻ của cùng một loài.
Một loại giao tiếp giữa các tế bào bằng tín hiệu phân tử được gọi là cảm biến số lượng (quorum sensing), phục vụ mục đích xác định liệu có mật độ đủ cao để đầu tư vào các quy trình chỉ thành công nếu số lượng lớn tương tự như trong việc bài tiết các enzym tiêu hóa hoặc phát ra ánh sáng.
Cảm biến số lượng cho phép vi khuẩn phối hợp biểu hiện gen và cho phép chúng sản xuất, giải phóng và phát hiện các chất tự động hoặc pheromone tích tụ với sự phát triển của quần thể tế bào.
Cây phát sinh loài (Classification seeks) thể hiện sự đa dạng của vi khuẩn so với các sinh vật khác. Ở đây vi khuẩn được đại diện bởi ba siêu nhóm chính: vi khuẩn siêu vi khuẩn CPR, vi khuẩn Terrabacteria và vi khuẩn Gracilicutes theo các phân tích bộ gen gần đây (2019).

Cây phát sinh loài (Classification seeks)
Phân loại tìm cách mô tả sự đa dạng của các loài vi khuẩn bằng cách đặt tên và phân nhóm các sinh vật dựa trên những điểm tương đồng. Vi khuẩn có thể được phân loại dựa trên cấu trúc tế bào, sự trao đổi chất của tế bào hoặc dựa trên sự khác biệt trong các thành phần tế bào, chẳng hạn như DNA, axit béo, sắc tố, kháng nguyên và quinon. Mặc dù các chương trình này cho phép xác định và phân loại các chủng vi khuẩn, nhưng vẫn chưa rõ liệu những khác biệt này thể hiện sự khác biệt giữa các loài riêng biệt hay giữa các chủng của cùng một loài. Sự không chắc chắn này là do thiếu các cấu trúc đặc biệt ở hầu hết các vi khuẩn, cũng như sự chuyển gen ngang (lateral gene) giữa các loài không liên quan. Do chuyển gen ngang, một số vi khuẩn có quan hệ họ hàng gần có thể có hình thái và chuyển hóa rất khác nhau. Để khắc phục tình trạng không chắc chắn này, phân loại vi khuẩn hiện đại nhấn mạnh vào hệ thống hóa phân tử, sử dụng các kỹ thuật di truyền như xác định tỷ lệ guanin cytosine, lai bộ gen-gen, cũng như xác định trình tự các gen chưa trải qua quá trình chuyển gen bên rộng rãi, chẳng hạn như gen rRNA. Việc phân loại vi khuẩn được xác định bằng cách công bố trên Tạp chí Quốc tế về Vi khuẩn có Hệ thống (Bergey International Journal of Systematic Bacteriology) và Sổ tay về Vi khuẩn có Hệ thống (Bergey's Manual of Systematic Bacteriology). Ủy ban quốc tế về vi khuẩn học có hệ thống - International Committee on Systematic Bacteriology (ICSB) duy trì các quy tắc quốc tế về việc đặt tên vi khuẩn và các phân loại phân loại cũng như xếp hạng chúng trong Bộ luật quốc tế về danh mục vi khuẩn (International Code of Nomenclature of Bacteria).
Trong lịch sử, vi khuẩn được coi là một phần của họ Thực vật, giới Thực vật, và được gọi là "Schizomycetes" (nấm phân hạch). Vì lý do này, vi khuẩn tập thể và các vi sinh vật khác trong vật chủ thường được gọi là "hệ thực vật" (flora). Thuật ngữ vi khuẩn theo truyền thống được áp dụng cho tất cả các sinh vật nhân sơ đơn bào, cực nhỏ. Tuy nhiên, hệ thống học phân tử cho thấy sự sống của sinh vật nhân sơ bao gồm hai lĩnh vực riêng biệt, ban đầu được gọi là Vi khuẩn Eubacteria và Cổ khuẩn, nhưng hiện nay được gọi là Vi khuẩn và Cổ khuẩn tiến hóa độc lập với tổ tiên chung cổ đại. Cổ khuẩn và sinh vật nhân thực có quan hệ gần gũi với nhau hơn là vi khuẩn. Hai miền này cùng với sinh vật nhân thực là cơ sở của hệ thống ba miền, hiện là hệ thống phân loại được sử dụng rộng rãi nhất trong vi sinh vật học. Tuy nhiên, do sự ra đời tương đối gần đây của hệ thống học phân tử và sự gia tăng nhanh chóng số lượng trình tự bộ gen sẵn có, việc phân loại vi khuẩn vẫn là một lĩnh vực đang thay đổi và mở rộng.

Phân loại vi khuẩn
Việc xác định vi khuẩn trong phòng thí nghiệm đặc biệt có liên quan trong y học, nơi việc điều trị chính xác được xác định bởi loài vi khuẩn gây nhiễm trùng. Do đó, nhu cầu xác định các tác nhân gây bệnh ở người là động lực chính cho sự phát triển của các kỹ thuật xác định vi khuẩn.
Nhuộm Gram được phát triển vào năm 1884 bởi Hans Christian Gram, đặc trưng cho vi khuẩn dựa trên đặc điểm cấu trúc của thành tế bào của chúng. Các lớp peptidoglycan dày trong thành tế bào Gram dương có màu tím, trong khi thành tế bào mỏng Gram âm có màu hồng. Bằng cách kết hợp hình thái học và nhuộm Gram, hầu hết vi khuẩn có thể được phân loại thuộc một trong bốn nhóm (cầu khuẩn Gram dương, trực khuẩn Gram dương, cầu khuẩn Gram âm và trực khuẩn Gram âm). Một số sinh vật được xác định tốt nhất bằng các vết bẩn khác với vết Gram, đặc biệt là vi khuẩn mycobacteria hoặc Nocardia, cho thấy tính bền axit trên Ziehl-Neelsenhoặc các vết bẩn tương tự. Các sinh vật khác có thể cần được xác định bằng sự phát triển của chúng trong môi trường đặc biệt, hoặc bằng các kỹ thuật khác, chẳng hạn như huyết thanh học (serology).
Kỹ thuật nuôi cấy được thiết kế để thúc đẩy sự phát triển và xác định các vi khuẩn cụ thể, đồng thời hạn chế sự phát triển của các vi khuẩn khác trong mẫu. Thường thì các kỹ thuật này được thiết kế cho các mẫu vật cụ thể. Ví dụ: một mẫu đờm sẽ được xử lý để xác định sinh vật gây viêm phổi, trong khi mẫu phân được nuôi cấy trên môi trường chọn lọc để xác định sinh vật gây tiêu chảy, đồng thời ngăn chặn sự phát triển của vi khuẩn không gây bệnh. Các mẫu vật thường vô trùng, chẳng hạn như máu, nước tiểu hoặc dịch tủy sống, được nuôi trong các điều kiện được thiết kế để phát triển tất cả các sinh vật có thể. Khi một sinh vật gây bệnh đã được phân lập, nó có thể được đặc trưng thêm bởi hình thái, các kiểu sinh trưởng (chẳng hạn như sinh trưởng hiếu khí hoặc kỵ khí), kiểu tán huyết và nhuộm màu.
Cũng như phân loại vi khuẩn, việc xác định vi khuẩn ngày càng được sử dụng nhiều hơn bằng các phương pháp phân tử (molecular) và quang phổ khối lượng (mass spectroscopy). Hầu hết vi khuẩn chưa được xác định đặc điểm và có thể có những loài không thể nuôi cấy trong phòng thí nghiệm. Chẩn đoán bằng cách sử dụng các công cụ dựa trên DNA, chẳng hạn như phản ứng chuỗi polymerase ngày càng phổ biến do tính đặc hiệu và tốc độ của chúng so với các phương pháp dựa trên nuôi cấy. Các phương pháp này cũng cho phép phát hiện và xác định các tế bào sống được nhưng không thể nuôi cấy đang hoạt động trao đổi chất nhưng không phân chia. Tuy nhiên, ngay cả khi sử dụng các phương pháp cải tiến này, tổng số loài vi khuẩn vẫn chưa được biết đến và thậm chí không thể ước tính một cách chắc chắn. Theo phân loại hiện nay, có ít hơn 9.300 loài sinh vật nhân sơ đã biết, bao gồm vi khuẩn và cổ khuẩn; nhưng những nỗ lực để ước tính số lượng thực sự của sự đa dạng vi khuẩn đã dao động từ 107 đến 109 và thậm chí những ước tính đa dạng này có thể bị sai lệch theo nhiều cấp độ.
Bất chấp sự đơn giản rõ ràng của chúng, vi khuẩn có thể hình thành các liên kết phức tạp với các sinh vật khác. Các mối quan hệ cộng sinh này có thể được chia thành chủ nghĩa ký sinh (parasitism), chủ nghĩa tương hỗ (mutualism) và chủ nghĩa tương sinh (commensalism).
Tương tác với các sinh vật khác
Từ "commensalism" có nguồn gốc từ từ "commensal", có nghĩa là "ăn cùng bàn" và tất cả thực vật và động vật đều bị vi khuẩn commensal xâm chiếm. Ở người và các động vật khác, hàng triệu con sống trên da, đường hô hấp, ruột và các lỗ thông khác. Được gọi là "hệ thực vật bình thường" hoặc "commensals", những vi khuẩn này thường không gây hại nhưng có thể xâm nhập vào các vị trí khác của cơ thể và gây nhiễm trùng. Escherichia coli là một sinh vật sống trong ruột người nhưng có thể gây nhiễm trùng đường tiết niệu. Tương tự, liên cầu khuẩn là một phần của hệ thực vật bình thường trong miệng người.
Một số loài vi khuẩn tiêu diệt và sau đó tiêu thụ các vi sinh vật khác, những loài này được gọi là vi khuẩn ăn thịt (predatory bacteria). Chúng bao gồm các sinh vật như Myxococcus xanthus, tạo thành các đám tế bào tiêu diệt và tiêu hóa bất kỳ vi khuẩn nào mà chúng gặp phải. Các vi khuẩn này tiêu thụ trực tiếp các vi khuẩn khác hoặc bám vào con mồi để tiêu hóa chúng và hấp thụ chất dinh dưỡng hoặc xâm nhập vào tế bào khác và nhân lên bên trong bào tương (cytosol). Những vi khuẩn săn mồi này được cho là đã tiến hóa từ các đại thực bào (saprophages) tiêu thụ các vi sinh vật đã chết, thông qua sự thích nghi cho phép chúng quấn lấy và giết chết các sinh vật khác.
Một số vi khuẩn hình thành các liên kết không gian chặt chẽ cần thiết cho sự tồn tại của chúng. Một sự liên kết tương hỗ như vậy được gọi là sự chuyển giao hydro giữa các loài, xảy ra giữa các cụm vi khuẩn kỵ khí tiêu thụ các axit hữu cơ, chẳng hạn như axit butyric hoặc axit propionic tạo ra hydro, và vi khuẩn methanogenic archaea tiêu thụ hydro. Các vi khuẩn trong nhóm này không thể tiêu thụ axit hữu cơ vì phản ứng này tạo ra hydro tích tụ trong môi trường xung quanh chúng. Chỉ có sự liên kết mật thiết với loài vi khuẩn tiêu thụ hydro mới giữ cho nồng độ hydro đủ thấp để vi khuẩn phát triển.
Trong đất, vi sinh vật cư trú ở tầng sinh quyển (rhizosphere - vùng bao gồm bề mặt rễ và lớp đất bám vào rễ sau khi lắc nhẹ) thực hiện quá trình cố định đạm, chuyển khí nitơ thành các hợp chất chứa nitơ. Điều này cung cấp một dạng nitơ dễ hấp thụ cho nhiều loài thực vật, chúng không thể tự cố định nitơ. Nhiều vi khuẩn khác được tìm thấy như là sinh vật sống cộng sinh ở người và các sinh vật khác. Ví dụ: sự hiện diện của hơn 1.000 loài vi khuẩn trong hệ thực vật đường ruột bình thường của con người có thể góp phần vào khả năng miễn dịch đường ruột, tổng hợp vitamin, chẳng hạn như axit folic, vitamin K và biotin, chuyển đổi đường thành axit lactic cũng như lên men carbohydrate phức tạp không tiêu hóa được. Sự hiện diện của vi khuẩn đường ruột này cũng ức chế sự phát triển của vi khuẩn có khả năng gây bệnh (thường thông qua loại trừ cạnh tranh) và những vi khuẩn có lợi này được bán dưới dạng thực phẩm chức năng bổ sung probiotic.
Gần như tất cả sự sống của động vật đều phụ thuộc vào vi khuẩn để tồn tại vì chỉ có vi khuẩn và một số cổ khuẩn mới sở hữu các gen và enzym cần thiết để tổng hợp vitamin B12, còn được gọi là cobalamin và cung cấp nó qua chuỗi thức ăn. Vitamin B12 là một loại vitamin tan trong nước, tham gia vào quá trình trao đổi chất của mọi tế bào trong cơ thể con người. Nó là một đồng yếu tố trong quá trình tổng hợp DNA, và trong cả quá trình chuyển hóa axit béo và axit amin. Nó đặc biệt quan trọng trong hoạt động bình thường của hệ thần kinh thông qua vai trò của nó trong tổng hợp myelin.
Cơ thể liên tục tiếp xúc với nhiều loài vi khuẩn, bao gồm cả vi khuẩn có lợi, phát triển trên da và màng nhầy, và vi khuẩn hoại sinh chủ yếu phát triển trong đất và vật chất thối rữa. Máu và dịch mô chứa các chất dinh dưỡng đủ để duy trì sự phát triển của nhiều vi khuẩn. Cơ thể có các cơ chế bảo vệ cho phép nó chống lại sự xâm nhập của vi sinh vật vào các mô của nó và tạo cho nó khả năng miễn dịch tự nhiên hoặc khả năng đề kháng bẩm sinh chống lại nhiều vi sinh vật. Không giống như một số vi rút, vi khuẩn tiến hóa tương đối chậm nên nhiều bệnh do vi khuẩn cũng xảy ra ở các động vật khác.
Nếu vi khuẩn hình thành mối liên kết ký sinh với các sinh vật khác thì chúng được xếp vào nhóm tác nhân gây bệnh. Vi khuẩn gây bệnh là nguyên nhân chính gây tử vong và bệnh tật ở người và gây ra các bệnh nhiễm trùng như uốn ván (do Clostridium tetani), sốt thương hàn, bạch hầu, giang mai, dịch tả, bệnh do thực phẩm, bệnh phong (do Micobacterium leprae) và bệnh lao (gây ra do Mycobacterium tuberculosis). Nguyên nhân gây bệnh cho một bệnh y khoa chỉ có thể được phát hiện nhiều năm sau đó, như trường hợp của Helicobacter pylori và bệnh loét dạ dày tá tràng. Các bệnh do vi khuẩn cũng xảy ra trong nông nghiệp, với vi khuẩn gây bệnh đốm lá, cháy lá và héo rũ trên cây trồng, cũng như bệnh Johne, viêm vú, vi khuẩn salmonella và bệnh than ở động vật trang trại.
Mỗi loài mầm bệnh có một phổ tương tác đặc trưng với vật chủ của chúng. Một số sinh vật, chẳng hạn như Staphylococcus hoặc Streptococcus có thể gây nhiễm trùng da, viêm phổi, viêm màng não và nhiễm trùng huyết, một phản ứng viêm toàn thân tạo ra sốc, giãn mạch lớn và tử vong. Tuy nhiên, những sinh vật này cũng là một phần của hệ vi sinh bình thường của con người và thường tồn tại trên da hoặc trong mũi mà không gây ra bất kỳ bệnh tật nào. Các sinh vật khác luôn gây bệnh cho người, chẳng hạn như Rickettsia, là ký sinh nội bào bắt buộc chỉ có thể phát triển và sinh sản trong tế bào của các sinh vật khác. Một loài Rickettsia gây ra bệnh sốt phát ban, trong khi một loài khác gây ra bệnh sốt đốm Rocky Mountain. Chlamydia, một nhóm ký sinh nội bào bắt buộc khác, chứa các loài có thể gây viêm phổi hoặc nhiễm trùng đường tiết niệu và có thể liên quan đến bệnh tim mạch vành. Một số loài, chẳng hạn như Pseudomonas aeruginosa, Burkholderia cenocepacia và Mycobacterium avium là mầm bệnh cơ hội và gây bệnh chủ yếu ở những người bịức chế miễn dịch hoặc xơ nang. Một số vi khuẩn tạo ra độc tố, là nguyên nhân gây bệnh. Đây là nội độc tố (endotoxins), đến từ các tế bào vi khuẩn bị hỏng và ngoại độc tố (exotoxins) được tạo ra bởi vi khuẩn và thải ra môi trường. Ví dụ: vi khuẩn Clostridium botulinum tạo ra extoxin cực mạnh gây tê liệt hô hấp và Salmonellae tạo ra nội độc tố gây viêm dạ dày ruột. Một số ngoại độc tố có thể được chuyển đổi thành độc tố, được sử dụng làm vắc-xin để ngăn ngừa bệnh.
Nhiễm trùng do vi khuẩn có thể được điều trị bằng thuốc kháng sinh, được phân loại là diệt khuẩn nếu chúng tiêu diệt vi khuẩn hoặc kìm khuẩn nếu chúng chỉ ngăn chặn sự phát triển của vi khuẩn. Có nhiều loại kháng sinh và mỗi loại ức chế một quá trình khác nhau đối với mầm bệnh được tìm thấy trong vật chủ. Một ví dụ về cách kháng sinh tạo ra độc tính chọn lọc là chloramphenicol và puromycin ức chế ribosome của vi khuẩn, nhưng không phải ribosome của sinh vật nhân thực khác nhau về cấu trúc. Thuốc kháng sinh được sử dụng cả trong điều trị bệnh cho người và nuôi thâm canh để thúc đẩy sự phát triển của động vật, nơi chúng có thể góp phần vào sự phát triển nhanh chóng của tình trạng kháng thuốc kháng sinh trong quần thể vi khuẩn. Có thể ngăn ngừa nhiễm trùng bằng các biện pháp sát trùng như khử trùng da trước khi dùng kim tiêm đâm vào da và bằng cách chăm sóc ống thông trong nhà đúng cách. Các dụng cụ phẫu thuật và nha khoa cũng được khử trùng để tránh bị nhiễm khuẩn. Các chất khử trùng như thuốc tẩy được sử dụng để tiêu diệt vi khuẩn hoặc các mầm bệnh khác trên bề mặt để ngăn ngừa ô nhiễm và giảm nguy cơ nhiễm trùng hơn nữa.
Vi khuẩn axit lactic như loài Lactobacillus và loài Lactococcus kết hợp với nấm men và nấm mốc, đã được sử dụng hàng ngàn năm để chế biến thực phẩm lên men, chẳng hạn như pho mát, dưa chua, nước tương, dưa bắp cải, giấm, rượu. và sữa chua.
Tầm quan trọng của vi khuẩn trong công nghệ và công nghiệp
Khả năng phân hủy nhiều loại hợp chất hữu cơ của vi khuẩn là rất đáng chú ý và đã được sử dụng trong xử lý chất thải và xử lý sinh học. Vi khuẩn có khả năng tiêu hóa hydrocacbon trong dầu mỏ thường được sử dụng để làm sạch dầu tràn. Phân bón đã được thêm vào một số bãi biển ở Prince William Sound trong nỗ lực thúc đẩy sự phát triển của những vi khuẩn tự nhiên này sau vụ tràn dầu Exxon Valdez năm 1989. Những nỗ lực này đã có hiệu quả trên những bãi biển không bị phủ quá dày bởi dầu. Vi khuẩn cũng được sử dụng để xử lý sinh học các chất thải độc hại trong công nghiệp. Trong ngành công nghiệp hóa chất, vi khuẩn quan trọng nhất trong việc sản xuất các hóa chất tinh khiết đối quang để sử dụng làm dược phẩm hoặc hóa chất nông nghiệp.
Vi khuẩn cũng có thể được sử dụng thay thế thuốc trừ sâu trong việc kiểm soát dịch hại sinh học. Điều này thường liên quan đến Bacillus thuringiensis (còn gọi là BT), một loại vi khuẩn Gram dương, sống trong đất. Các phân loài của vi khuẩn này được sử dụng làm thuốc trừ sâu đặc hiệu Lepidopteran dưới các tên thương mại như Dipel và Thuricide. Vì tính đặc hiệu của chúng, những loại thuốc trừ sâu này được coi là thân thiện với môi trường, ít hoặc không ảnh hưởng đến con người, động vật hoang dã, động vật thụ phấn và hầu hết các loài côn trùng có ích khác.
Do khả năng phát triển nhanh chóng và khả năng thao tác tương đối dễ dàng, vi khuẩn là đối tượng nghiên cứu cho các lĩnh vực sinh học phân tử, di truyền và hóa sinh. Bằng cách tạo đột biến trong DNA của vi khuẩn và kiểm tra các kiểu hình thu được, các nhà khoa học có thể xác định chức năng của gen, enzym và con đường trao đổi chất ở vi khuẩn, sau đó áp dụng kiến thức này cho các sinh vật phức tạp hơn. Mục đích hiểu được hóa sinh của tế bào đạt đến biểu hiện phức tạp nhất của nó trong việc tổng hợp một lượng lớn dữ liệu động học và biểu hiện gen của enzyme thành các mô hình toán học của toàn bộ sinh vật. Điều này có thể đạt được ở một số vi khuẩn được nghiên cứu kỹ lưỡng, với các mô hình chuyển hóa Escherichia coli hiện đang được sản xuất và thử nghiệm. Sự hiểu biết này về sự trao đổi chất và di truyền của vi khuẩn cho phép sử dụng công nghệ sinh học đối với vi khuẩn kỹ thuật sinh học để sản xuất các protein điều trị, chẳng hạn như insulin, các yếu tố tăng trưởng hoặc kháng thể.
Vì tầm quan trọng của chúng đối với nghiên cứu nói chung, các mẫu của các chủng vi khuẩn được phân lập và bảo quản trong các Trung tâm Tài nguyên Sinh vật (Biological Resource Centers). Điều này đảm bảo sự sẵn có của chủng này cho các nhà khoa học trên toàn thế giới.
Vi khuẩn lần đầu tiên được quan sát bởi nhà hiển vi người Hà Lan Antonie van Leeuwenhoek vào năm 1676 bằng cách sử dụng kính hiển vi một thấu kính do chính ông thiết kế. Sau đó, ông đã công bố những quan sát của mình trong một loạt các bức thư gửi tới Hiệp hội Hoàng gia London (Royal Society of London). Vi khuẩn là khám phá hiển vi đáng chú ý nhất của Leeuwenhoek. Chúng chỉ ở mức giới hạn những gì thấu kính đơn giản của ông có thể tạo ra và trong một trong những thời điểm gián đoạn nổi bật nhất trong lịch sử khoa học, không ai khác có thể nhìn thấy chúng lần nữa trong hơn một thế kỷ. Các quan sát của ông cũng bao gồm các động vật nguyên sinh mà ông gọi là “animalcules” và phát hiện của ông đã được xem xét lại dưới những phát hiện gần đây hơn về lý thuyết tế bào.
Christian Gottfried Ehrenberg đưa ra từ "vi khuẩn" vào năm 1828. Trên thực tế, sinh vật mà ông tìm ra là một chi chứa vi khuẩn hình que không sinh bào tử, trái ngược với Bacillus - một chi hình que sinh bào tử. Vi khuẩn có hình dạng được Ehrenberg xác định năm 1835.
Louis Pasteur đã chứng minh vào năm 1859 rằng sự phát triển của vi sinh vật gây ra quá trình lên men và sự phát triển này không phải do tự phát (nấm men và nấm mốc, thường liên quan đến quá trình lên men, không phải là vi khuẩn, mà là nấm). Cùng với Robert Koch đương thời, Pasteur là người sớm ủng hộ lý thuyết vi trùng của bệnh tật. Trước họ, Ignaz Semmelweis và Joseph Lister đã nhận ra tầm quan trọng của việc khử trùng tay trong y tế. Ý tưởng của Semmelweis đã bị từ chối và cuốn sách của ông về chủ đề này bị cộng đồng y tế lên án, nhưng sau đó các bác sĩ của Lister đã bắt đầu vệ sinh tay của họ vào những năm 1870. Trong khi Semmelweis, người bắt đầu với các quy tắc về rửa tay trong bệnh viện của mình vào những năm 1840 trước khi lan truyền ý tưởng về chính vi trùng và cho rằng các bệnh là do "phân hủy chất hữu cơ động vật".

Louis Pasteur đã chứng minh vào năm 1859 rằng sự phát triển của vi sinh vật
Robert Koch, một nhà tiên phong trong lĩnh vực vi sinh y học, đã nghiên cứu về bệnh tả, bệnh than và bệnh lao. Trong nghiên cứu của mình về bệnh lao, Koch cuối cùng đã chứng minh được lý thuyết về vi trùng, mà ông đã nhận được giải Nobel vào năm 1905. Trong các định đề của Koch, ông đã đưa ra các tiêu chí để kiểm tra xem một sinh vật có phải là nguyên nhân gây ra bệnh hay không, và các định đề này vẫn còn được sử dụng ngày nay.
Ferdinand Cohn được cho là người sáng lập ngành vi khuẩn học, nghiên cứu vi khuẩn từ năm 1870. Cohn là người đầu tiên phân loại vi khuẩn dựa trên hình thái của chúng.
Mặc dù vào thế kỷ 19, người ta biết rằng vi khuẩn là nguyên nhân của nhiều bệnh, nhưng vẫn chưa có phương pháp điều trị kháng khuẩn hiệu quả nào. Năm 1910, Paul Ehrlich đã phát triển loại thuốc kháng sinh đầu tiên, bằng cách thay đổi thuốc nhuộm chọn lọc Treponema pallidum - loại xoắn khuẩn gây bệnh giang mai - các hợp chất tiêu diệt mầm bệnh một cách có chọn lọc. Ehrlich đã được trao giải Nobel năm 1908 cho công trình nghiên cứu miễn dịch học và đi tiên phong trong việc sử dụng chất nhuộm màu để phát hiện và xác định vi khuẩn, với công trình nghiên cứu của ông là cơ sở của phương pháp nhuộm Gram và Ziehl-Neelsen.
Một bước tiến lớn trong nghiên cứu vi khuẩn đến vào năm 1977 khi Carl Woese nhận ra rằng cổ khuẩn có một dòng tiến hóa riêng biệt từ vi khuẩn. Phân loại phát sinh loài mới này phụ thuộc vào trình tự sắp xếp của RNA ribosome 16S, và chia sinh vật nhân sơ thành hai miền tiến hóa, như một phần của hệ thống ba miền.
Tin bài khác









 nhiệm kỳ 2025-2030")


![[Video] Đại hội nhiệm kỳ V Hội các Phòng thử nghiệm Việt Nam (VINALAB) nhiệm kỳ 2025-2030](/resize/363/tin-hoi-vien/dai-hoi-dai-bieu-toan-quoc-lan-thu-5-221225-1x363x4.jpg)